
Bob Cowen during the poster presentation where he outlined the concept of planktonportal
Bob Cowen and Jessica Luo got to present the concept of the Plankton portal to the oceanography community at the 2014 Ocean Science Meeting in Honolulu, Hawaii!

Bob Cowen during the poster presentation where he outlined the concept of planktonportal
Bob Cowen and Jessica Luo got to present the concept of the Plankton portal to the oceanography community at the 2014 Ocean Science Meeting in Honolulu, Hawaii!
You may think orcas or great white sharks are the most voracious predators in the oceans, but based on their abundance and ability to consume a wide range of prey items, chaetognaths (a.k.a. “arrow worms”) give those big animals a run for their money. Large predators like sharks are extremely rare, but scoop up a bucket of seawater almost anywhere in the world and you are likely to find a few chaetognaths (if you have a microscope handy). Chaetognaths are transparent worms that often remain motionless in the water column, apparently relying on the element of surprise to capture a wide variety of plankton, including copepods, appendicularians, small fish larvae, and smaller chaetognaths. Chaetognaths are thought to be generalist feeders because their stomach contents often reflect the community captured by plankton nets. They use a mass of chitinous hooks around their mouths to capture prey – which gives them their name (“chaetognath” translates from Latin to mean “hairy jaw”) and a notoriously menacing appearance.

Chaetognaths are often straight in the ISIIS images but can also swim rapidly for short distances. The camera typically cannot resolve the tiny chitinous hooks on the chaetognath’s mouth.
Chaetognaths comprise about 100 species that are all typically 1-2 cm long. They are most abundant along the coasts, with some species being so sensitive to salinity that oceanographers can identify discrete water masses based solely on the community of chaetognath species. Similar to many other types of zooplankton, chaetognaths are hermaphrodites, first being male then changing into female as they get larger. Fertilized eggs can be attached to vegetation or encased in a gelatinous web. Eggs then hatch into juvenile chaetognaths, and thus they have no larval stage. This is called direct development because there is no process of metamorphosis.

A clear image of the chaetognath’s mouth on the cover of Current Biology.
The chaetognath’s body is streamlined and adapted to feeding with minimal visual input. The have sensory cilia that can detect small vibrations in the water that tell the chaetognaths that prey is within striking distance. With a quick flick of its tail, the chaetognath surges forward to capture the prey in its chitinous hooks used for grasping. It then transfers the prey to its mouth where it is swallowed whole. Some deeper water chaetognaths (>700 m deep) can even use bioluminescence to create a cloud of light that scientists think can be used to escape predation (Haddock and Case 1994).

The most handsome chaetognath found by our citizen scientists!
References:
Haddock SHD and Case JF (1994) A bioluminescent chaetognath. Nature 367:225
Johnson WS and Allen DM (2005) Zooplankton of the Atlantic and Gulf coasts: A guide to the identification and ecology. Johns Hopkins University Press, Baltimore, MD
Lalli CM and Parsons TR (1997) Biological oceanography an introduction. Elsevier Butterworth-Heinemann, Burlington, MA

Artwork by Jean-Olivier Irisson
Great news! We are working on translating Plankton Portal in French with our
French Collaborators: Fabrice Not from the Station Biologique de Roscoff and Jean-Olivier Irisson from the Observatoire Océanologique and Station Zoologique de Villefranche-sur-mer. “The idea is also to get some interest from French schools to develop a curriculum around Plankton Portal” Dr. Irisson explains. Stay Tuned.
Of all plankton groups, probably most is known about the copepods. They represent a critical link in the food chain and are consumed by diverse animal community ranging in size from small fish, chaetognaths, and ctenophores all the way up to large whales (the right whale is a voracious copepod feeder). Because of their small size and importance as food, copepods are affectionately known as “the rice of the sea.” Copepods are effectively captured by plankton nets because they have hard exoskeletons, and scientists have good estimates of their abundances and distributions. Although copepods are all relatively small (0.5 mm – 5 mm in length), they comprise over 200 families and 10,000 different species.

Examples of typical copepods. Note to the two large appendages on the top of the head with small sensory hairs
Copepods consume both phytoplankton and microzooplankton in two different ways: suspension feeding and raptorial feeding. Suspension feeding is relatively passive and performed by beating small appendages that draw a current through a feeding chamber. Copepods then select which particles encountered are food and discard others. Raptorial feeding is used to actively capture prey. Many copepods have small sensors on their first appendages to detect water disturbances produced by prey and also predators. They can use these relatively large appendages to “hop” through the water and capture an unsuspecting prey item or to quickly escape a predator.
Copepod reproductive strategies vary greatly and are adapted towards the ability to withstand the variable conditions that characterize the ocean environment. For example, many copepod eggs have the ability to enter a phase of diapause where they remain viable on the bottom for several months or even years, only hatching with conditions are favorable (high concentrations of food). Some copepods carry their eggs, allowing them to develop a bit before releasing them into the water column. The timing of copepod reproduction is especially important for the life cycle of fishes because most fish larvae depend on the recently hatched copepod nauplii for food. If there are not enough copepod nauplii present when fish larvae are abundant, there could be mass starvation events causing few fish larvae to reach their juvenile stage. Because of this, the copepod life cycle is extremely important to fish populations and overall ocean ecosystem health.

This image was taken from a thin layer near Stellwagen Bank offshore of Massachusetts, USA. Each one of the white particles is a copepod. The concentration of organisms in this image corresponds to ~400,000 individuals per cubic meter! That is some good eating for a right whale!
One of the most remarkable characteristics of copepods is their tendency to aggregate in discrete thin layers within the water column. Sometimes >90% of the copepod biomass will be confined these thin layers, which are a maximum of 5 m thick. ISIIS and other systems that sample on small scales are ideal for detecting these layers of copepods, and the function of the formation and dissipation of copepod thin layers is not well understood. Copepods have been shown to be attracted to strong changes in current direction and speed, potentially allowing them to feed at a faster rate within these zones (Woodson et al. 2005). The changes in environmental variables associated with aggregations of copepods are of great interest to marine ecologists. With your help, we can better understand how these extremely important organisms are distributed throughout our oceans!
References:
Johnson WS, Allen DM (2005) Zooplankton of the Atlantic and Gulf coasts: A guide to their identification and ecology. Johns Hopkins University Press. Baltimore, MD.
Woodson CB, Webster DR, Weissburg MJ, Yen J (2005) Response of copepods to physical gradients associated with structure in the ocean. Limnol Oceanogr 50:1552-1564
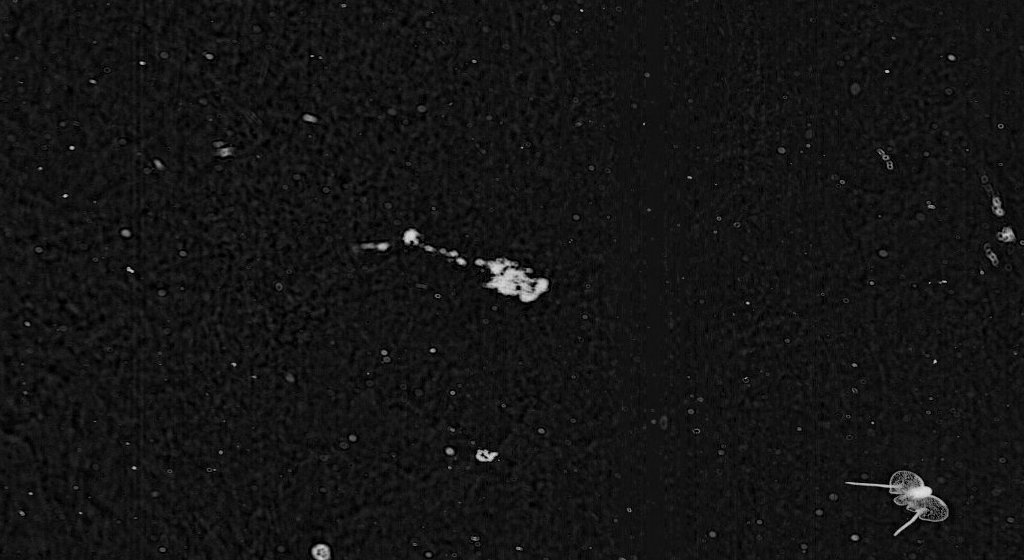
Pteropods are a group of organisms that we’re not focusing on because they are not very abundant in the Plankton Portal dataset. Nevertheless, you may have run across a few of those fascinating little creatures.
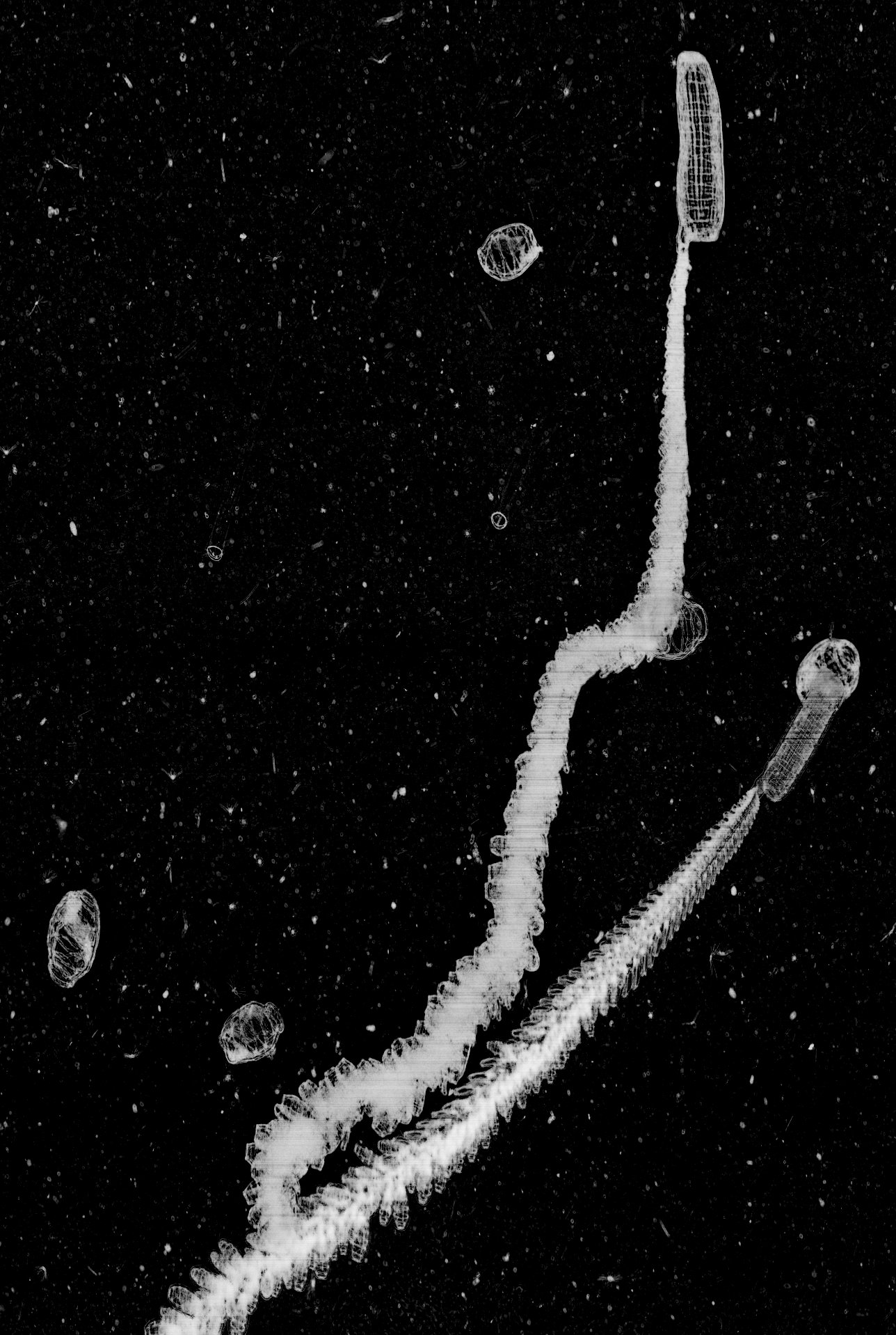
Pteropod, which means ‘wing-foot’ in Greek, is a group of free-swimming pelagic gastropods (snails). Officially, the word ‘pteropod’ is no longer used in taxonomy; it is a collective term which refers to two clades of gastropods—thecosome (shelled body) and gymnosome (naked body). Pteropods are quite unique because in order to adapt to life in the water column, their foot is modified into two wing-like flippers used for swimming. Their body size ranges from a few millimeters to several centimeters – so they’re easily imaged by ISIIS. They can be quite abundant in certain regions of the world’s oceans, and are typically found near surface waters.
The first group of pteropods, thecosomes, are also known as the sea butterflies. They have a pair of large ‘wings’ and swims by continually flapping them. Their body is encased in a delicate and translucent shell.The shell can be coiled, needle-like, triangular, and globed.

Thecosomes are omnivores. Their diet consists of diatoms, dinoflagellates, and zooplanktons such as copepods, tintinnids, and other gastropod larvae. They capture food by secreting a spherical mucus web several times larger than their body. Scientists believe that the use of the large size mucus web is to capture large, fast swimming prey, such as copepods. The web acts as a filter: particles that are too large for ingestion are removed. During feeding, the mucus web is suspended above the animal while the animal remains motionless below. Ciliary action draws back the web to the mouth and the whole web is ingested.

Thecosome reproductive biology is quite unusual. The animal first matures and functions as male. The male pteropod mates with another male and the sperm is stored until the animal changes into a female. When the animal turns into female; its male reproductive organs degenerate. The female lays fertilized floating egg mass that later hatch into swimming larvae (veliger).
When a thecosome dies, its shell sinks to the bottom of the sea and forms sediment called pteropod ooze. The shell is composed of aragonite, an unstable form of carbonate mineral. Anthropogenic ocean acidification is one of the challenges that pteropods face. The increase of anthropogenic carbon dioxide level in the atmosphere reduces pH and carbonate ion concentration in the ocean, thus decreasing the calcium carbonate saturation level. As a result, the production of biogenic carbonate becomes more difficult. Overall, they have a hard time secreting their protective shell because of ocean acidification.
The second group of pteropods, or gymnosomes, are more commonly known as sea angels. They have much smaller wings which appear as side lobes. They are more robust and lack a shell. Unlike their thecosome relatives, gymnosomes are carnivores. They are active hunters and exclusively prey on thecosome pteropods. A combination of hooks and a toothed radula are employed to extract the flesh from the thecosomes’ shells.
The reproductive anatomy of gymnosome pteropods is similar to thecosomes pteropods. The only difference: the male reproductive organs do not degenerate in females. Gymnosomes has two distinct larvae forms. Eggs are hatched into shelled veliger. The veliger metamorphoses into a shell-less polytrochous larvae. The polytrochous larvae are initially wingless and movement is achieve by three ciliary bands. They gradually grow wings and lose the ciliary bands as they become adults.
Here is a very nice video about Pteropods.
Plankton Chronicles Project by Christian Sardet, CNRS / Noe Sardet and Sharif Mirshak, Parafilms. See Plankton Chronicles interactive site: planktonchronicles.org
Salps and doliolids (class Thaliacea) are interesting animals because they are in the phylum Chordata, which includes all animals with a notochord during development (e.g., humans, fish, cats), but thaliaceans have a vastly different appearance and feeding strategy compared to most vertebrates. A salp or doliolid body is essentially a giant pumping muscle that forces water through a mucous net filter that collects phytoplankton and is ingested periodically. Both groups have limited mobility, with salps using muscular contractions to scoot through the water, while doliolids use tiny beating cilia to propel themselves.

A salp in the process of forming a new chain of clones for asexual reproduction (see white coil)
The life history of salps and doliolids is remarkable and complex. Similar to plants, their life cycle alternates between sexual and asexual generations. The solitary phase reproduces asexually by budding off clones of itself. On salps, a chain of these clones develops on the solitary animal that is then released and reproduces sexually with other salp chains. The chains first mature as female and then change sexes to become male when they are larger! These chains release small solitary salps that then begin asexual budding once they are a certain size. Doliolids on the other hand produce short-lived tadpole larvae that are not seen in salps. When you consider that a chain of salps contains an average of ~28 individuals, it is no surprise that these organisms are capable of extremely fast reproductive rates and can double their populations in hours (Heron 1972). Some scientists think their remarkable reproductive rates can overwhelm other phytoplankton grazers, which could explain the fact that large salp aggregations are often associated with low biomass of other grazers (Alldredge and Madin 1982).
Doliolids imaged offshore of Monterey Bay showing asexual budding
Because of their ability to reproduce quickly, salps are often very abundant near steady supplies of phytoplankton, such as at ocean fronts (zones where two water masses with differing physical properties meet) and eddies (Deibel and Paffenhöfer 2009). However, these organisms cannot tolerate extremely dense aggregations of phytoplankton because their mucous filters will become clogged with prey, which severely decreases their feeding efficiency. Salps and doliolids can “bloom” like other jellies, and when these blooms die off the dead salp bodies can export a large amount of carbon into deeper waters. Because of salps and doliolids close evolutionary relationship to vertebrates, scientists are also very interested in their developmental biology. Scientists are trying to use salps as a model organism to study the development of complex nervous systems in all vertebrate animals (Lacalli and Holland 1998).
Check out this video from Plankton Chronicles on these remarkable animals!
Plankton Chronicles Project by Christian Sardet, CNRS / Noe Sardet and Sharif Mirshak, Parafilms. See Plankton Chronicles interactive site: planktonchronicles.org
References:
Alldredge AL and Madin LP (1982) Pelagic tunicates: Unique herbivores in the marine plankton. Bioscience 32:655-663
Deibel D and Paffenhöfer GA (2009) Predictability of patches of neritic salps and doliolids (tunicata, thaliacea). J Plankton Res 31:1571-1579
Heron AC (1972) Population ecology of a colonizing species: The pelagic tunicate Thalia democratica – I. individual growth rate and generation time. Oecologia 10:269-293
Lacalli TC and Holland LZ (1998) The developing dorsal ganglion of the salp Thalia democratica, and the nature of the ancestral chordate brain. Philosophical Transactions of the Royal Society B: Biological Sciences 353:1943-1967
As you may know by now, the In Situ Ichthyoplankton Imaging system (ISIIS) is the instrument we used to obtain the images of plankton for Plankton Portal. It captures images as it travels through the water column. The idea behind this imaging system was to film a large volume of water in order to detect and image relatively rare zooplankton, like larval fish and small jellies.
So why are the images are black and white, very contrasted and almost like a line drawing? Our challenge in designing this instrument was to be able to employ macro-photography at a fast speed, obtain a large depth of field (large volume sampled), while minimizing motion blur. After researching for a few months we settled on an imaging technique that could answer our demands: Shadow imaging or focus shadowgraphy!
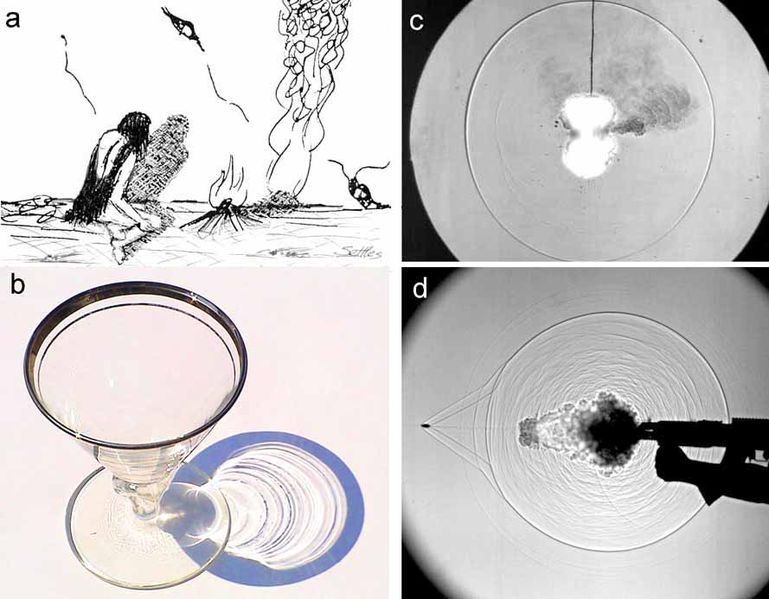
a) prehistoric shadowgraphy, b) sunlight shadowgram of a martini glass, c) “focused” shadowgram of a common firecracker explosion, d) “Edgerton” shadowgram of the firing of an AK-47 assault rifle
Gary S. Settles
This technique is actually not new and was used extensively for the study of shockwaves as well as ballistic. The idea is to cast a shadow onto a sensor or film instead of trying to directly record the imaged object. Let me explain: since most plankton are small and quite transparent, imaging using a traditional camera must rely on ambient sunlight. In this scenario, you won’t see much because the organisms blend into the surrounding water. Imaging a shadow cast by ISIIS reveals their distinct shape and location. It’s like looking at the shadows at the bottom of a swimming pool created by the sun going through the water! In this case, we do not use the ambient sunlight, but create our own light beam using a blue LED light and a set of mirror and lenses. The light is collimated, meaning that the light beam travels in a tight, parallel direction like that of a laser thus ensuring that even over long distances, we can create a very good shadow. We then use a specific set of lenses aligned with the camera to capture this shadowgraph image. Since the light beam is directed toward the camera sensor, it allows for very high speed imaging and avoids motion blur when moving through the water. Lastly, we invert the images for aesthetic purposes on the site, and voila! Now you have a beautiful set of black and white images of plankton for the world to see!
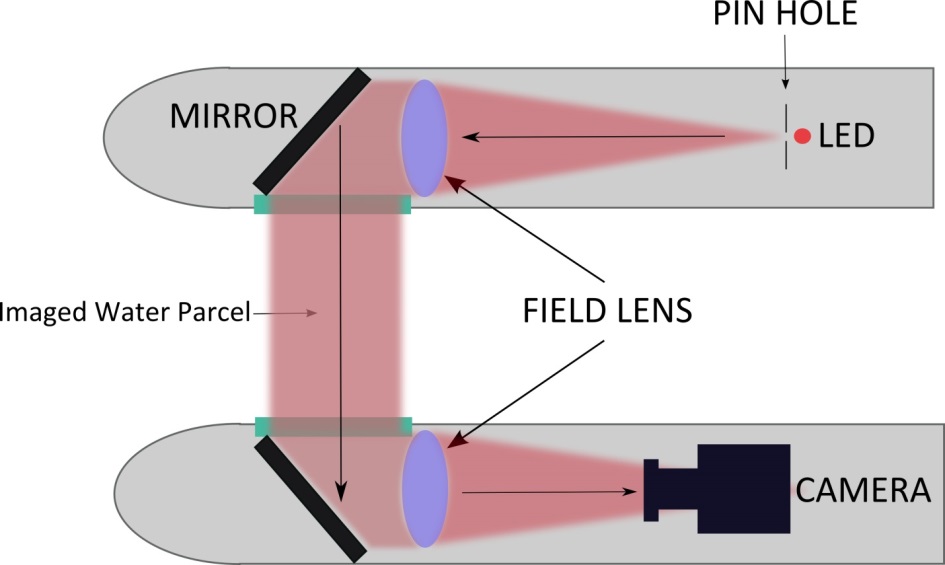
This schematic shows the optical alignment inside ISIIS. A bean of light is collimated by a large lens and the refocused after going through the water. The camera records the shadows casted by the plankton as the moves behind the ship
So for all the people who asked about why some of the ctenophores (like lobates, beroids, and cydippids) were so ‘overexposed,’ now you know. These animals are dense and not very transparent, thus casting a hard shadow onto the lens. Instead of appearing black as a typical shadow, we have inverted the image and now they appear white.

Lobate ctenophore

Since the 1800’s, plankton has been studied and collected using simple nets with very fine mesh. The process of analysis of these plankton ‘samples’ is tedious, labor intensive, confined to laboratory, and can only be done on relatively small areas of the ocean. By sampling in this manner, it is difficult to get a good understanding of how planktonic organisms are distributed and how they interact with each other. Yet plankton represents a very important part of a global system feeding larger animals like fish, whales and many others. The In Situ Ichthyoplankton Imaging System (ISIIS) is one of a few systems in the world capable of improving the way we study plankton to better understand their life and function in the marine environment. Instead of using an actual net to capture plankton, ISIIS captures the images of the organisms and information about their immediate surroundings. ISIIS samples continuously, resulting in a collection of digital images that record the exact location of the various plankton organisms in relation to each other and the environment in which they live. Further, the images are recorded onto a simple hard drive instead of slurry of plankton all mixed together in a sample jar with formaldehyde (yech!).

ISIIS is an underwater imaging system developed to capture real time images of plankton that are relatively rare, small, and fragile such as fish larvae and delicate gelatinous organisms (like jelly fish). ISIIS is composed not only of a macro-camera system with its own illumination but it also is integrated into an underwater vehicle with a variety of additional sensors to measure the depth, salinity and temperature of the water, as well as such properties as dissolved oxygen, light level, and even how much chlorophyll a (measure of primary production) is present. Together, the camera and sensors provide detailed profiles and tracks of what plankton are where and what the ocean environment around them is like.
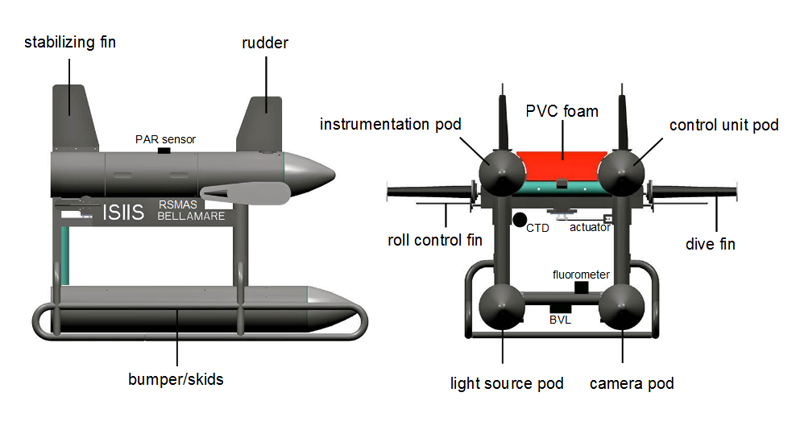
The vehicle, and associated imaging system and sensors, moves up and down through the water column using side-mounted, user-controlled dive fins (like an underwater glider) while being towed behind an oceanographic ship moving at 5 knots. The vehicle frame is divided into four compartmentalized enclosures with imaging and optical equipment seamlessly integrated into ISIIS’s ventral housings and environmental sensors and electronics in the dorsal housings. ISIIS is designed to undulate in a zigzag fashion between the surface and a maximum depth of 200 meters.

The ISIIS system utilizes imaging technology very similar to an office scanner flipped on its side. The imaged parcel of water passes between the forward portions of two streamlined pods where it is “scanned” and transformed into a continuous image. The resulting very high-resolution image is of plankton in their natural position and orientation. When a sufficient volume of water is imaged this way, quantification of concentration (individuals per unit volume) and fine scale distribution is possible. ISIIS is capable of imaging a maximum of 162 Liters (43 gallons) of water per second (when moving at 5 knots) with a pixel resolution of 70 µm (the thickness of a human hair).
The imaging data and associated oceanographic data are sent to the surface ship via a fiber optic cable and recorder onto a main computer for later viewing and analysis.

