The Science Team is currently out in the field in the Straits of Florida, on the R/V Walton Smith, sampling with both ISIIS and MOCNESS (Multiple Opening Closing Net and Environmental Sampling System), on an 18-day cruise titled OSTRICH (Observations on Subtropical TRophodynamics of ICHthyoplankton).
The overall goal of this NSF-sponsored project is to quantify the patterns and consequences of the fine-scale to sub-mesoscale distributions of larval fishes, their prey, and their predators near and across a major western boundary current passing through the Straits of Florida. By sampling a series of water masses at very high resolution, this study addresses specific hypotheses concerning: i) the drivers of aggregations and patchiness, and ii) the biological consequences of predator-prey interactions at fine scales.
Sample ISIIS images showing diversity of plankton from multiple coastal sites (including the Southern California Bight!)
Sampling involves a novel combination of detailed in situ sampling of the horizontal and vertical distributions of plankton, targeted fine-scale net sampling, and analyses of individual-level recent daily larval growth to enable the identification of the biological and physical processes driving fine-scale plankton distributions.
Follow along on the ISIIS facebook page as we periodically post updates (via our terrible internet connection at sea!) and also check out this cool video made by one of our cruise participants, Chris Muiña:
The name for these worms literally means “many bristles,” which refers to the “legs” that they use to move through the water. These surprisingly fast animals are predators of copepods, appendicularians, and even small larval fishes. Most polychaetes are meroplankton, meaning that they are plankton only for their egg and larval stages. When they reach a certain size, they settle out of the water column and spend their adult lives associated with some kind of substrate (e.g., reefs, sand, mud, rock, etc.). A few species are holoplanktonic, spending their entire lives drifting in the ocean currents.
One genus of holoplanktonic polychaete that we have encountered in the ISIIS images is Tomopteris. These 2-5 cm polychaetes feature several adaptations that are favorable for life in water column. First, they are highly transparent, allowing them to blend into the surrounding water. If they could be easily seen, polychaetes would be tasty little snacks for fish. One thing that limits their transparency is a gut that runs down the middle of the body. When this gut is full, the polychaete is easier to see because it cannot hide a stomach full of food! Someone might hypothesize that polychaetes that have recently eaten might be more susceptible to visual predators, such as fish, but to date, no one has explored this question. Second, their “legs” have paddle-shaped ends with two lobes, which improve their swimming ability compared to other groups of polychaetes (Todd et al. 1996).
Many of these polychaetes (Tomopteris spp.) are actively swimming. The gut runs down the center of the animal between the legs, but it is difficult to see in these images.
Although polychaetes are relatively rare plankton, we did manage to see a good number of them near Stellwagen Bank, Massachusetts, USA. The graph shows the vertical distribution of the Tomopteris polychaetes along two ISIIS transects. As you can see, Tomopteris polychaetes were predominantly found in deeper waters. In the images taken, it is difficult to see the gut, which would show up as a white line running down the middle of the body. This means that these individuals had not eaten recently, so what are they doing in the deep waters? Possibly hiding from predators in waters with less light? Or could this behavior be related to mating? Only with further research can we find out what influences the distributions of these and other planktonic animals.
Near Stellwagen Bank, Massachusetts, USA, many of the polychaetes tended to reside deeper in the water column. They are virtually absent from the top 20 m.
Unidentified polychaete larvae imaged by ISIIS in the Gulf of Mexico.
References:
Todd CD, Laverack MS, Boxshall GA (1996) Coastal Marine Zooplankton: A practical manual for students (2nd ed.) Cambridge University Press, New York.
As organisms that cannot swim against the currents, plankton are intimately connected to their physical environment. Many species are quite sensitive to the temperature, salinity, and nutrient levels that either lead to their proliferation or demise. Physical conditions and nutrient levels can lead to high abundances of particular plankton types. These plankton “blooms” are common throughout the world’s oceans and can be composed of phytoplankton, zooplankton, or gelatinous zooplankton, depending on the environmental conditions.
Generally phytoplankton (plankton that use photosynthesis like plants) need nutrients and light to grow at very high rates. Since light is readily available in the surface ocean, nutrient availability is the most important driver of phytoplankton blooms. Phytoplankton blooms of most concern to environmental monitoring groups are often described as Harmful Algal Blooms (HABs). Some HABs composed of diatom species Pseudo-nitzschia spp. or the dinoflagellate Karenia breviscan produce toxins harmful to copepods, fish, and higher trophic levels like dolphins and humans. “Red tides” are actually blooms of Karenia brevis that sometimes lead to massive fish dieoffs. Other phytoplankton blooms are harmful not because of the toxins that they produce, but because of the processes that happen when the blooms die off: massive amounts of phytoplankton die and sink to the bottom where they are decomposed by bacteria. These bacteria use oxygen to consume the dead phytoplankton, creating large portions of the water column that are low in oxygen. Fishes and some zooplankton avoid these low oxygen zones, but gelatinous zooplankton seem to be able to withstand low oxygen conditions. These low oxygen regions are often referred to as “dead zones” because very few animals can live there. A dead zone occurs regularly in the summertime in the northern Gulf of Mexico and has been expanding in recent years. Reducing nutrient/fertilizer runoff from farmlands and cities is therefore crucial to limiting the growth of phytoplankton and maintaining healthy coastal ecosystems.
Blooms of zooplankton can form via two different mechanisms 1) currents from different water masses merge to create a dense patch of organisms, or 2) consistently favorable conditions allow the zooplankton to reproduce faster than their predators can consume them. These two mechanisms are distinguished in the scientific literature as “apparent blooms” and “true blooms” (Graham et al. 2001). Apparent blooms can result from converging currents such as fronts, or the behavior of zooplankton aggregating along some kind of physical discontinuity, such as a thermocline. True blooms are typically the result of high food concentrations, high survival of larvae/juveniles, or a combination of multiple factors. For scientists, it is sometimes difficult to know what mechanism led to a zooplankton bloom, and they need to consider the history of the water masses where the zooplankton are found to figure out how the bloom formed (Greer et al. 2013). Zooplankton blooms associated with copepods are generally considered to be healthy for the ecosystem. The timing of copepod peak abundances with the first feeding of larval fishes is thought to be an important factor contributing to the variation in fish population abundances (Cushing 1975). Blooms of jellyfish, on the other hand, are often associated with ecosystems that are environmentally degraded through high nutrient input or consistent overfishing (Jackson et al. 2001), but there are some scientists who think jelly blooms are simply a characteristic of their life histories (Condon et al. 2013). More data on jellyfish abundances over longer time periods will help scientists understand relationships between the environment and the frequency of jellyfish blooms. You can also report jellyfish sightings at http://www.jellywatch.org.
Here is avideo of one ISIIS downcast through the water column of near a frontal feature offshore of San Diego, CA, USA. It shows a bloom of Solmaris spp. jellies concentrated near the surface. Convergent currents that are commonly seen at fronts likely contributed to the formation of this bloom.
References:
Condon RH, Duarte CM, Pitt KA, Robinson KL, Lucas CH, Sutherland KR, Mianzan HW, Bogeberg M, Purcell JE, Decker MB, and others (2013) Recurrent jellyfish blooms are a consequence of global oscillations. Proc Natl Acad Sci U S A 110:1000-1005
Cushing DH (1975) Marine ecology and fisheries. Cambridge University Press, London
Graham WM, Pagès F, Hamner WM (2001) A physical context for gelatinous zooplankton aggregations: A review. Hydrobiologia 451:199-212
Greer AT, Cowen RK, Guigand CM, McManus MA, Sevadjian JC, Timmerman AHV (2013) Relationships between phytoplankton thin layers and the fine-scale vertical distributions of two trophic levels of zooplankton. Journal of Plankton Research 35:939-956
Jackson JBC, Kirby MX, Berger WH, Bjorndal KA, Botsford LW, Bourque BJ, Bradbury RH, Cooke R, Erlandson J, Estes JA, and others (2001) Historical overfishing and the recent collapse of coastal ecosystems. Science 293:629-637
Here is a nice collection of plankton underwater shot by Dr. Evan D’alessandro while conducting research offshore Miami
the Portuguese Man of War is not a Jellyfish but a Siphonophore!
The Portuguese Man of War or Physalia is a siphonophore floating at the surface of the ocean like a drifting balloon with deadly tentacles waiting to capture careless preys. Its body looks like a sail that catches the ocean wind to propel itself.
The Portuguese Man of War Fish or Nomeid seems to be immune to the powerful sting of the Physalia. It is actually a very agile swimmer that can avoid the stinging tentacles.It uses this deadly siphonophore for shelter against predators and can also feed on some of the smaller tentacles that do not seems to have a strong sting.
Nice ctenophore
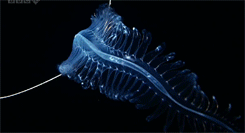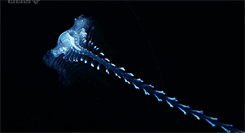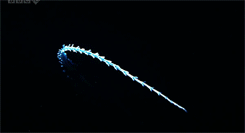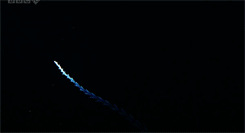
Our favorite, the Venus Belt! Nice to see this one in color!
salp chain
If you click on the this amazing picture you will see a small fish larvae seeking shelter among the salps. very neat!
ctenophore bloom
Beautiful bloom of lobate ctenohphores. The wall of death some smaller plankters:)
Bob Cowen during the poster presentation where he outlined the concept of planktonportal
Bob Cowen and Jessica Luo got to present the concept of the Plankton portal to the oceanography community at the 2014 Ocean Science Meeting in Honolulu, Hawaii!
Hey plankton hunters! This week we are showing off four exceptional zooplankton found by you, our keen-eyed and inquisitive citizen scientists. We are amazed at how many plankton species have been uncovered on the site and just how capable you all have been at discerning some truly tricky taxa from the varying forms and shapes captured by the ISIIS camera. We thank all of the citizen scientists for your participation on the Plankton Portal! These images found by our citizen scientists continue to excite and we are eager to discover what resides in the thousands of images yet to be seen by human eyes!
This capture of an anthomedusa is definitely a prime example of how the images captured by ISIIS can be equal measures fine-resolution biological data and one-of-a-kind organismal artwork. This gelatinous organism is baring all for us in this frame and we get a clear view of not only the striations on the exterior of the bell (the exumbrella) unique to this species and the fully extended tentacles, but also the central gastric pouch (stomach) appearing as the dark mass within the bell and the internal network of radial canals where digested food is transported. I think I can also see this critter blushing as ISIIS takes the snapshot! This medusa shown here is relatively uncommon in the images provided for you from the Southern California Bight, and we couldn’t be happier that our fantastic and dedicated group of citizen scientists spotted this gelatinous beauty. Annatiara affinis is a hydromedusa like many of the #4tentacles and #morethanfourtentacle medusae found on the site. The unique (and photogenic) lines appearing along the exterior of the bell were very helpful in pinning down an ID for this critter. From what we have seen, this seems to be a rare image captured of Annatiara where the tentacles are fully extended from the margin of the bell, and we are extremely grateful that this lovely jelly was so at ease in front of the ISIIS cam.
Shrimp — #Shrimp
This is one of the largest shrimps I have seen on the portal and provides a great side-view of the crustacean anatomy. The orientation of this shrimp with the abdomen tucked under the carapace (upper shell) and the antenna trailing sharply away from the head indicates that it is moving rapidly towards its posterior (bottom left of image), using a swimming stroke known as the “cardioid escape reaction”—slapping the abdomen shut and quickly propelling the crustacean away from the perceived danger. This specific behavior played an important role in the field of neuroscience, in fact. When it was discovered, this behavioral response was the first example of a “command neuron mediated behavior”— meaning a specific behavioral pattern resulting from the stimulation of a single neuron. I wonder what stimulated this crustacean’s command neuron? Perhaps it is camera shy.
Arrow Worm / Chaetognath — #ArrowWorm
I’m curious if that dark blob may be some out-of-luck plankter soon to be nabbed by this voracious predator. I am especially fond of these in focus captures of chaetognaths. The dart-shaped body and the hydrodynamic taper of the paired lateralfins really show off the sleek and elegant body plan of these brutal invertebrate carnivores. The chaetognath body has a protective outer covering known as a cuticle, a tough but flexible non-mineral layer exterior to the epidermis. Chaetognaths are notoriously efficient predators and hunt other planktonic organisms using hooked grasping spines that flank the mouth. A hood arising from the neck region can be drawn over or away from the hunting spines, much like the action of sheathing and unsheathing a blade. Equipped with an armor of cuticle and sword-like spines these guys are definitely well suited for combat!
Physonect Siphonophore — #Sipho #Corncob
The siphonophores love to put on a good show for us here on the portal and this frame is truly exceptional. The shadowgraph imaging technique used by ISIIS lends itself to capturing in detail the elaborate gelatinous structures displayed by these colonial organisms. Siphonophores are comprised of many single animals, or zooids, which are highly specialized and coordinated in function. The zooids of a physonect siphonophore arise from a long stem at the end of which is a gas-filled float referred to as a pneumatophore. The pneumatophore is on display in this image here appearing as the dark, oval-shaped appendage on the upper left end of the main “body.” The portion that resembles a corn on the cob is referred to as the nectosome. The nectosome is composed of many swimming bells, or nectophores, each one of which is a single medusoid zooid. These nectophores display remarkable coordination among each other and the selective contraction of these zooids allows for the siphonophore to move and turn in any and all directions. Physonect siphonophores are predators and rely on long, branching tentacles for prey capture. The one whipping across the frame here is definitely on the prowl. Each tentacle arises from a single feeding polyp situated below the nectosome in a region called the siphosome. You can see the siphosomal region on this specimen as the narrowing, darkly filled feature curling upward from the base of the nectosome. They sure have a lot of ‘somes’ and ‘phores’ but we forgive their repetitive nomenclature because we are always glad to find some siphonophores.
We hope this has been a fun and informative look at a few of the many tremendous critters captured by ISIIS and found by the citizen scientists. If you come across an image you think is particularly cool on the portal then tag it with #FFF and we will check it out for use on the blog. As always, looking forward to the next Fantastic Find Fridays!
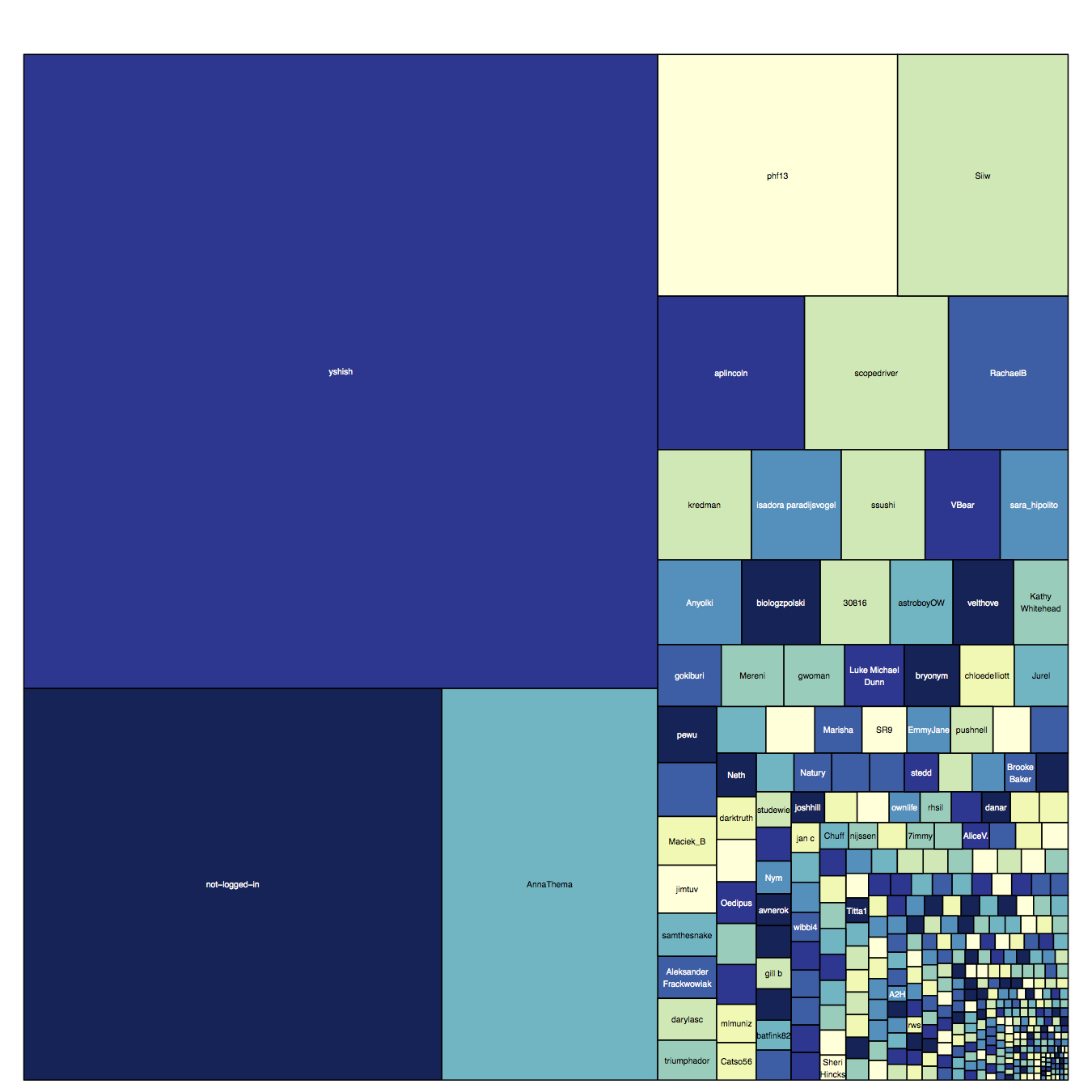
Thanks everyone for your phenomenal response to the treemap that I posted for the 300,000 classifications post. It’s really neat to be able to see how different users have classified more, or less over time. Of course, if you are one of the top classifiers, it’s fun to see your name up there!
At the suggestion of Lee Henderson, MD (wow! cool to see people from all professions and walks of life on Plankton Portal), I made an updated treemap with classifications from the past month. If there’s interest in periodic updates in this manner, let me know in the comments and I can post one of these up every couple weeks. But I don’t want to make it too much of a competition. Actually, it probably is a giant competition. So compete away!
Top 10 classifiers from the past month:
1. yshish
2. AnnaThema
3. phf13
4. Siiw
5. aplincoln
6. scopedriver
7. RachaelB
8. kredman
9. isadora paradijsvogel
10. ssushi
Thanks everyone!
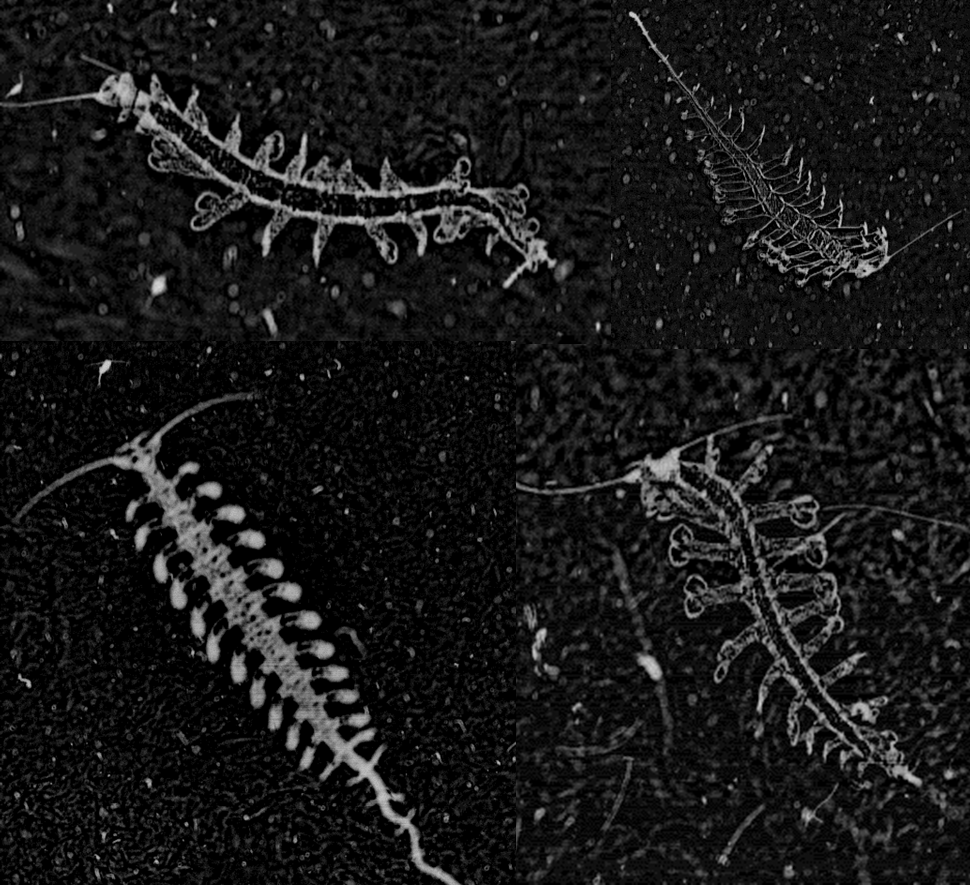
On another note, our star moderator yshish emailed me today with a few questions about some cool siphonophores she found on PP. (side note: yshish has the BEST image collections on Talk. I regularly will be able to find specific images or taxa that I’m looking for by going through her collections.) Anyway, check out this series of images:
You might remember that we featured the top image in a previous Fantastic Finds Friday (FFF) post. Now we bring it back because of the other two images that were found (the last one found 1 hr ago). These images found three months apart might be the same organism but in neighboring frames. We can’t tell right away, but once we go back into the raw data, we will be able to pull out the locations and times of these three images to check.
Now, the interesting question posed here is — is this siphonophore budding? Are these siphonophores asexually reproducing as we imaged it? And the answer — yes! absolutely! (I addressed this question earlier in a Talk post but now’s the time to feature it in the blogs!) All the little round bells that you see on the tentacles are small siphonophores developing from the gonozooids (reproductive ‘organs’) that will eventually be released and form free-swimming “eudoxids.” When they are released, these eudoxids will develop and then be capable of sexual reproduction. Weird, right? But wait, there’s more.
These eudoxids then develop little reproductive organs along its stem. Instead of eudoxids functionally male or female, they actually develop male and female reproductive parts – eggs and sperm – alternating between the two, sometimes regularly, sometimes irregularly. That way, they can ensure that the eggs are fertilized and can develop into larvae, then post-larvae, then adults.
The open ocean is a vast place, and animals have developed vastly different strategies for how to ensure the continuation of their species, whether it is in spawning aggregations (e.g. Grouper fish spawning in the Caribbean – and larvaceans also apparently form spawning aggregations) or being hermaphroditic (like Ctenophores and some fish), and being able to asexually bud and reproduce sexually. These siphonophores have adopted the strategy of being both hermaphroditic AND able to reproduce sexually and asexually.
This is the best diagram I’ve found to describe the life history of a Calycophoran siphonophore (all of the ‘rocketship’ and ‘two-cup’ siphonophores). This one is from C. Carre and D. Carre (1991).























{kind=link}